国内外辣椒抗黄瓜花叶病毒的研究总结,黄瓜花叶病毒能治愈吗
国内外辣椒抗黄瓜花叶病毒的研究总结
黄瓜花叶病毒能治愈吗?编辑1、加强产地运输和检疫,严格控制疫区封锁。加强对非疫区的检疫,防止种子和幼苗传播。葫芦科植物从国内发生区运往非发生区的种子和幼苗必须附有“植物检疫证书”,以确保无此类病毒,种植区应加强产地检查。

黄瓜花叶病毒在田间主要靠什么传播
病毒主要在多年生根植物上越冬。鸭跖草、反枝苋、刺梨、酸浆等。蚜虫是传播毒素的蚜虫的越冬宿主,如桃蚜和棉蚜,它们在甜瓜春季萌发后开始移动或迁移,成为疾病传播的主要媒介。 感染的子叶在苗期变黄并枯萎,幼叶呈深绿色和浅绿色叶状花。同时,病叶在不同程度上表现出萎缩和畸形。 成年植物被感染的新叶是黄色和绿色的,开有叶状的花,患病的叶子小而干瘪,叶子变厚,严重时叶子会卷回来。茎的节间缩短,茎变形,在严重的情况下,患病的叶子枯萎。瓜条呈现深绿色和浅绿色交替出现。1.高温和干旱引起的根部感染 2.蚜虫感染 3.叉子感染 黄瓜花叶病毒病的症状:可以发生在苗期到成虫期,但主要发生在成虫期。苗期感染了这种疾病,子叶变黄枯萎,幼叶呈现绿叶,深绿色和浅绿色交替出现。 成人疾病的发病始于上部新叶和褪绿斑的症状。后来,它的特点是厚而绿色的叶子和叶子斑驳。患病的叶子变得更小,略微萎缩。严重的叶缘是生物化学家王英来。在过去的半个世纪里,在营养、维生素、血红蛋白、酶和物质代谢方面取得了一系列重要成就。在担任中国科学院生物化学研究所所长和中国生物化学学会主席期间,他在该所的建设和学会的发展中发挥了重要作用。他是世界上第一个人。
黄瓜花叶病毒能治愈吗
黄瓜花叶病毒能治愈吗?编辑1、加强产地运输和检疫,严格控制疫区封锁。加强对非疫区的检疫,防止种子和幼苗传播。葫芦科植物从国内发生区运往非发生区的种子和幼苗必须附有“植物检疫证书”,以确保无此类病毒,种植区应加强产地检查。
黄瓜花叶病毒在田间主要靠什么传播
国内外辣椒抗黄瓜花叶病毒的研究总结范文

辣椒。属于茄科辣椒属,是中国第二大蔬菜作物。近年来,病害的流行已成为制约[胡椒生产的主要因素1]
。黄瓜花叶病毒(CMV)可引起严重的全身症状,导致叶片发黄、扭曲、严重的花叶,并损害水果的商业特性[2 -6】。黄瓜花叶病毒病一般会导致辣椒减产20%~ 30%,严重的情况下甚至在某些地区减产50%~ 60%。这是胡椒[最严重的疾病之一。黄瓜花叶病毒的检出率是我国辣椒产量最高的,也是辣椒抗病育种的主要目标国内外巨细胞病毒菌株鉴定和划分的基本方法是从不同种和植物中收集巨细胞病毒分离株,接种到不同种和属的不同宿主上,根据症状进行菌株划分,[19 -23]。这种划分方法不能反映病毒致病基因和宿主抗病基因之间的对应关系,在抗病育种中意义不大9级分类是[“八五”期间辣椒抗病育种重点研究组制定的分类标准,0级:无症状;一级:心脏叶明脉或接种叶的急性小枯萎病;第三级:系统镶嵌或茎上的坏死斑点;5级:茎上系统重马赛克、畸形或坏死条纹;7级:大多数叶片变形,蕨类植物叶片和植株矮小或茎、枝和脉系统坏死;9级:植物严重矮化,停止生长或遭受严重的系统性坏死,直到整株植物死亡[8]。该分级标准广泛应用于辣椒抗巨细胞病毒材料筛选、遗传规律分析和抗性育种中,[33,42,45,55,58]。此外,根据不同的指标还有其他分级标准。。一些研究者也提出了基因菌株的概念。希望CMV的毒力和抗病性能够联系在一起,为根据植物中局部病斑的数量划分疾病等级:植物中局部病斑的数量为0 ~ 100,等级1为0 ~ 5;二级为6 ~20级;三级为21 ~ 50;4年级50多[47】。根据酶联免疫吸附试验检测的感染百分比,制定分级标准:免疫(一),检出率为0;抗病(r),检出率为0。1% ~ 10%;中度抗体的检出率为10。1%~30%。中等灵敏度,30。1% ~ 50%;感受性,50。1%~ 100%[6,59]。的抗病育种提供有力的理论依据。然而,巨细胞病毒与品种之间的基因对基因关系仍不确定,品种抗性与巨细胞病毒毒力之间的一一对应也不清楚[26 -27]。由于巨细胞病毒株系划分没有统一的标准,建立一套通用的株系识别宿主谱是深入研究巨细胞病毒株系划分的前提。因此,从1987年到1988年,中国辣椒(甜椒)特殊群体专门建立了一套适合辣椒抗赤霉病育种的赤霉病鉴别寄主谱4抗黄瓜花叶病毒辣椒种质资源现状及抗性机制分析。利用这套不同的宿主谱,中国采集的20株巨细胞病毒优势分离株被分为6个菌株,即轻症(ProM)、环斑(PRi)、斑驳(PoM)、蕨叶(PF)、坏死(PN)和黄斑(PyM)种质资源是作物遗传育种的物质基础,也是抗病育种的重要前提。世界上三大辣椒种质资源是:世界蔬菜中心(原名亚洲蔬菜研发中心,AVRDC)是世界上最大的辣椒种质资源,包括11个辣椒品种,共8665份材料,占全球辣椒种质资源[60的11%;根据种质资源信息网(GRIN),美国国家植物种质中心有3411份辣椒种质资源。哥斯达黎加的热带农学研究和高等教育中心(cat-ie)收集了七种辣椒品种。。。传统育种方法耗时长,品种替换远远落后于病毒株的变异。分子标记辅助选择育种可以有效克服传统育种的缺陷,加快育种进程。目前,还没有抗巨细胞病毒辅助选择育种的分子标记。辣椒基因组数据的公布为抗巨细胞病毒研究提供了新的机会。因此,本研究对国内外辣椒抗黄瓜花叶病毒的研究进行了综述,提出了存在的问题和发展方向,为今后辣椒抗黄瓜花叶病毒的研究提供了一定的理论参考。
黄瓜花叶病毒简介
1.1黄瓜花叶病毒基因组
1916年,杜利特尔和贾格尔分别在[的密执安州和纽约州的黄瓜上发现了一种病毒,即黄瓜花叶病毒。巨细胞病毒是溴病毒科黄瓜花叶病毒的典型成员。巨细胞病毒是一种三联病毒,由三个直径约为28纳米的球形颗粒组成。基因组由三个基因组(RNA1: ~ 3 350 bp,RNA2: ~ 3 050 bp,RNA3: ~ 2 200 bp)和两个亚基因组(RN4A:~ 689 BP和RNA4: ~ 1 031 bp)组成,分别位于RNA2和RNA3的3’端。共有5个开放阅读框架(ORFs) [11-12]。由核糖核酸1编码的1a蛋白是病毒复制酶;由核糖核酸2编码的2a蛋白是病毒聚合酶。RNA4A翻译产生2b蛋白,其可影响病毒的全身感染并抑制病毒介导的基因沉默[13-14];由核糖核酸3的5’ORF编码的3a蛋白是驱动蛋白。3’端由核糖核酸4编码的蛋白质是病毒外壳蛋白[15 -17]。根据病毒生物学、血清学和分子特征,巨细胞病毒分离株可分为两个亚组(ⅰ和ⅱ),其中ⅰ亚组进一步分为ⅰa和ⅰb,ⅰ和ⅱ亚组的核酸序列同源性为69% ~ 77%,ⅰa和ⅰb的序列同源性大于95% 辣椒中巨细胞病毒感染的严重程度取决于感染时的苗龄。因此,适宜的苗龄对抗性鉴定非常重要。种子嫁接的苗龄一般包括亚叶期[6,36],1-2叶期[2,4,37 -38],3-4叶期[33,35,39 -45]和5-6叶期[7,46 -53]。综上所述,在国外研究中,接种期从子叶期到6叶期分布。在国内研究中,接种期主要集中在3-4叶期和5-6叶期。相关研究认为,苗期(子叶期和1 ~2叶期)过早,抗性鉴定材料的病害指数增加;幼苗过老(7 ~ 8片叶)时,抗病鉴定材料的病害指数下降。3 ~6叶期是种子嫁接的最佳时期,材料的抗性水平可以准确测定[54 -55]。。
1.2黄瓜花叶病毒株系的划分
[8]
黄瓜花叶病毒病的危害及症状
温带是巨细胞病毒的主要分布区,热带呈逐年增加的趋势,到目前为止,还没有发现辣椒材料对巨细胞病毒具有完全抗性,但是一些栽培辣椒的野生菌株和相关野生种对巨细胞病毒具有部分抗性。1994年,舒雅中心公布了辣椒抗病毒种质资源目录,包括165份抗巨细胞病毒材料和46份抗巨细胞病毒材料,广泛分布于[辣椒的5个栽培种、1个野生种和多个品种中。其中,一年生辣椒中发现113个抗黄瓜花叶病毒品种和41个抗黄瓜花叶病毒品种;灌木辣椒的野生种中,29种抗巨细胞病毒,13种抗巨细胞病毒,[61 -62]。。迄今为止,巨细胞病毒在100个科中寄生了1200多种植物,对各种蔬菜作物和观赏植物尤其有害,[30 -31]。巨细胞病毒主要通过各种蚜虫以不可持续的方式传播,一些植物可以通过种子传播病毒[18]。
寄主范围广、蚜虫传播和作物品种变化频率低,使巨细胞病毒每周都能存活,使其难以控制[3,31]。我国辣椒主产区巨细胞病毒检出率很高,北京、陕西、湖广和江浙地区检出率均在50%以上,[4、21、23、32]。
辣椒巨细胞病毒感染呈逐渐发生的过程。发病初期,嫩叶基部黄化,黄化症状逐渐延伸至整叶。然后新生的叶子显示出明显的马赛克。随着植物的发展,叶片出现严重萎缩,植株矮小,叶片颜色为暗深绿色[4,21,23,30,32。辣椒中巨细胞病毒感染的严重程度取决于感染时植株的年龄。一般来说,苗期感染巨细胞病毒会导致秃顶、矮化和减产,严重的话,还可能导致灭绝。成年病毒株的巨细胞病毒感染症状相对较轻,[2,33]。
辣椒对黄瓜花叶病毒的抗性鉴定
抗病性鉴定是抗病材料选择、抗病遗传规律研究和抗病品种选育的基础。关键是要准确反映材料的电阻水平。辣椒抗巨细胞病毒鉴定大多采用人工接种鉴定的方法。基本步骤包括接种液的配制、适宜苗龄的接种植株的选择、接种方法的选择和植株抗性的鉴定。
3.1接种溶液的制备
接种物的制备基本上遵循了1967年马拉鲁·[发明的方法,只是在不同的试验中略有改进。取纯化的巨细胞病毒病毒源,在烟草上繁殖,人工接种,接种后12 ~ 15天采集病叶。5ml 0.03摩尔/升磷酸盐缓冲液(Na2HPO4含0。2%二乙基二硫代氨基甲酸钠)加入到冰浴匀浆中的每1 g患病叶子中。滤液或上清液是接种物抗性机制主要是研究病毒与宿主的相互作用。抗性机理的定义是更好地利用抗性材料的基础。,用双层纱布过滤或以3 500转/分钟离心15分钟。
3.2接种植物苗龄的选择
[18]
几种辣椒对巨细胞病毒的抗性属于成虫抗性。苗期人工接种和鉴定很难发现其抗性,成虫接种难度更大。因此,有必要改变苗期常规接种方法,采用打顶接种方法。打顶接种是指在5叶期去掉幼苗的生长点,留下4片叶子,4天后人工揉搓第三片叶子。然后研究了第3、4叶轴侧枝的发生率。根据侧枝的发生率,可以判断植物的抗病性,从而可以表达材料的成虫抗性,从而提高人工接种鉴定CMV抗性[54,56]的准确性。
3.3疫苗接种方法的选择
巨细胞病毒人工接种方法包括搓指接种、磨砂玻璃勺接种、喷枪接种和病毒浸汁接种[35]。接种前,前三种方法需要在叶片表面喷洒一薄层(直径约23微米)碳化硅,接种后30分钟清洗多余的接种物。搓指接种和磨砂玻璃勺接种操作简单,但接种不均匀。在辣椒抗黄瓜花叶病毒遗传规律的研究中,手指摩擦接种法[2,4 -7,33,36 -37,49,57]最常用于抗病材料的选择和抗病育种。用于喷枪接种的喷枪为油漆喷枪,接种时喷枪距离幼苗约2厘米,空空压机工作压力为2。1 ~2。5 kg/cm2,喷枪接种适用于大量材料的抗性鉴定。该方法已部分应用于我国[35、39、54辣椒抗巨细胞病毒材料的筛选。病毒汁浸根法是指清洗幼苗根部的土壤,浸泡在接种液中。这种方法的接种效果最好,但工作量大,不容易操作,而且[使用不多。
3.4植物抗性鉴定标准
辣椒对巨细胞病毒的抗性主要根据病害分类标准进行分类。疾病分类标准主要包括3、5和9个分类标准。第三种分类是基于叶片镶嵌和收缩程度。0级:无症状;一级:轻微马赛克,无收缩;二级:重度马赛克,轻度收缩;等级3:重马赛克,严重收缩[2】。5级根据疾病的严重程度分类:0级:无症状;1级:局部坏死斑点或轻微失绿,无全身感染;二级:轻度至中度斑驳或马赛克,无明显叶片损伤;3级:中度至重度马赛克及明显的叶片畸形;四级:花叶严重,叶片明显萎蔫畸形;等级5:马赛克非常严重,叶子明显变形,黄化枯萎[38】。
[24]
[25]
[26]
4.1抗黄瓜花叶病毒辣椒种质资源现状
[28]
[29]
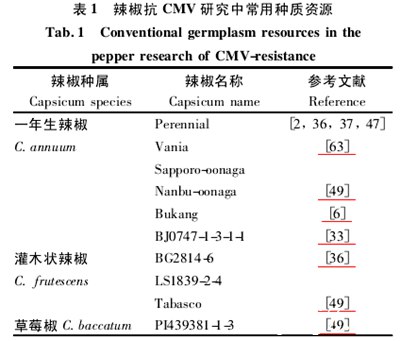
后来,研究人员在辣椒研究和育种中发现了一些新的抗巨细胞病毒材料,包括一年生辣椒(C. an-nuum)、灌木辣椒(C. frutescens)和草莓辣椒(c.baccatum)。具体信息如表1所示。
经过数百年的培育、驯化和选择,我国辣椒后代在不同的生态条件下形成了不同形态的辣椒种质资源。据2003年统计,已有2119份辣椒种质进入国家种质资源库。在“七五”和“八五”期间,我国研究人员鉴定了3000多份辣椒种质和新组合对疫病、花叶病毒和花叶病毒两种病害的抗性,筛选出一批单抗多抗优质种质[8]。随后,我国不断加强对辣椒抗巨细胞病毒种质的鉴定,并取得了一定的成果(表2)。
综上所述,世界上抗黄瓜花叶病毒的辣椒种质主要是一年生辣椒和灌木辣椒,用于抗性研究的材料相对单一。根据国内研究,962份鉴定材料中,免疫材料5份,高抗性材料2份,占0。5%和0。分别占总数的2%。以上结果表明,辣椒免疫和高抗性的黄瓜花叶病毒种质资源在国际和国内都非常稀缺,继续发现、鉴定和创造更多的新种质资源仍然是黄瓜花叶病毒抗性研究的重点。
4.2辣椒抗巨细胞病毒机理分析
[35]
目前,辣椒抗或耐巨细胞病毒的主要机制有三种。
抑制病毒侵入宿主细胞:研究表明,接种巨细胞病毒后多年生抗病材料的叶表面病斑数量明显少于易感材料约洛·旺德·[47,68];病毒繁殖的抑制:接种巨细胞病毒后,巨细胞病毒在多年生植物中的繁殖速率远低于易感材料约洛·旺德(Yolo Wonder)和另一种抗病材料瓦尼亚·[69];病毒运动的抑制:杜福尔等人[56]发现,CMV在接种易感物质Yolo Wonder后2天从接种的叶片迁移到叶柄,3天后出现全身症状。接种抗性材料米洛德和香草5天后,巨细胞病毒从接种的叶片迁移到叶柄,没有全身症状。辜莞允-沃迪姆等人[56,69]发现,与易感材料约洛·旺德(Yolo Wonder)相比,巨细胞病毒在抗性材料米洛德和瓦尼亚的传播延迟了4-6周,而巨细胞病毒的发病率和感染率明显较低。
上述结果表明,不同的电阻材料具有不同的电阻机理,而同一种材料可能同时具有两种或两种以上的电阻机理。辣椒抗巨细胞病毒的遗传分析。
辣椒抗巨细胞病毒遗传分析是利用抗病基因提高栽培品种抗病性的前提,国内外对此进行了大量研究。关于辣椒抗巨细胞病毒遗传,世界上主要有三种观点。前期科研结果表明,辣椒对黄瓜花叶病毒的抗性受单基因控制,表现为隐性遗传[70-72];Kang等人([6)最近的研究表明,安农杆菌对布卡菌(Bukang)对巨细胞病毒的抗性由一个基因控制,表现出显性遗传。Pochard等人,[73]认为辣椒材料Rama对巨细胞病毒的抗性是由具有数量效应的显性基因Riv控制的。随后,许多研究证实辣椒对巨细胞病毒的抗性受多个基因控制,表现出部分显性遗传[2,33,37,47,63]。
相对而言,国内对辣椒抗巨细胞病毒遗传的研究结果基本一致:辣椒抗巨细胞病毒遗传符合“加性-显性”模式,加性效应更为重要;抗性遗传是不完全显性遗传。抗性是由多个基因控制的数量性状。广义遗传力和狭义遗传力都很高。研究内容详见表3。
辣椒抗巨细胞病毒的QTL作图及连锁标记分析
6.1辣椒抗巨细胞病毒的QTL作图
国内外大多数研究认为辣椒对巨细胞病毒的抗性是一个数量性状,同时定位了多个与巨细胞病毒抗性相关的QTL位点。QTL分析表明,主要的数量性状基因很少,大多数是次要的数量性状基因。QTL之间存在互动效应。与其相连的分子标记的遗传距离相对较大,在目前的研究水平上,这些标记不能充分用于辣椒抗巨细胞病毒育种。相关结果列于表3。
6.辣椒抗巨细胞病毒相关基因的2个连锁标记分析
辣椒抗巨细胞病毒连锁标记的研究很少,仅报道了2例。吴小莉·[7]利用通过CMV抗性材料VC16a和敏感材料SS69杂交获得的F2群体,选择了与辣椒CMV抗性位点相连的ISSR标记ⅰ-34,遗传距离为27。3 cM。Kang等人[6]命名了安奴姆对布卡克姆1号的抗性基因。利用同源序列比对和BAC文库筛选方法,开发了三个与其相连的单核苷酸多态性标记,即CaTm-int3HRM、CaT1616BAC和240H02sp6,遗传距离为2 cM。目前,辣椒抗巨细胞病毒育种尚无分子标记。
辣椒抗巨细胞病毒研究的7个问题及发展方向
辣椒抗巨细胞病毒的研究已经做了大量的工作,取得了一定的进展,但还没有取得突破性进展,这表明以往的研究存在一些问题,需要采取相应的措施逐步解决。总结如下:缺乏免疫和高抗巨细胞病毒资源。从综述中可以看出,辣椒免疫和高抗巨细胞病毒种质资源非常匮乏,导致辣椒抗巨细胞病毒育种缺乏后续研究。辣椒种质资源的发现、鉴定、改良和创新仍然是辣椒抗巨细胞病毒研究的重点。收集、鉴定和利用野生辣椒种质资源,利用种间杂交等技术创造种间杂种,拓宽辣椒遗传背景是获得抗巨细胞病毒种质资源的有效方法。抗性机制的不确定性及抗性材料的遗传规律。
巨细胞病毒的不同分离株、病毒株的差异以及病毒的重组和变异都可能导致不同抗性材料中的多重抗性机制和不同遗传规律[76]。辣椒抗巨细胞病毒机制的研究仅限于发病时期、病毒数量等表型分析,抗病材料的抗性机制在分子水平上尚未阐明。经典遗传方法只能用于评价整个拮抗遗传规律,不能分析单个基因的遗传效应。这需要深化病毒与宿主相互作用的研究,在分子水平上阐明抗性机制和抗性遗传规律,更好地利用抗性材料。分子标记很少。目前,RFLP、RAPD、SSR等标记主要用于辣椒抗巨细胞病毒的分子遗传学研究。由于辣椒遗传基础狭窄,亲本间多态性标记较少,这些标记在应用分析群体中不能均匀覆盖整个基因组。基于此,推测通过应用具有相对较远关系的组合或应用新类型的标记,例如Indel和SNP,可以在空白色区域中发现新的抗性位点。辣椒基因组信息[77]的公布为SNP和Indel标记的开发和应用提供了可能性,并能有效解决辣椒分子标记缺乏的问题。
基因定位效率低。在辣椒基因组序列公布之前,辣椒相关基因作图主要采用传统的基因作图方法,如作图克隆和分离群体分离分析(BSA),需要开发大量的分子标记来构建遗传图谱,工作量大,精度低。辣椒基因组数据的公布不仅可以大规模开发序列特异性分子标记,为分子标记辅助育种提供条件,也为基于基因组数据的基因定位技术的应用奠定了必要的条件[78,79]。安倍[80]提出的多重图谱方法(MutMap method)类似于QTL序列原理,即在混合池后对分离群体中特定表型植物的基因进行重新测序。然而,MutMap主要针对突变基因,QTL序列可以通过分析重测序数据的单核苷酸突变来分析数量性状和定位目标基因。这两种方法工作量小、效率高、精度高,是新一代基因定位技术。因此,辣椒基因组数据和新一代基因定位技术应在辣椒抗巨细胞病毒研究中得到充分利用,以提高辣椒抗巨细胞病毒基因定位效率。
参考:
[1]克罗斯比·K·M·蔬菜ⅱ·[·M]。柏林:斯普林格,2008:221 - 248。
[2][2]查姆·阿·布,格鲁贝·罗·C,拉皮多特·M,等.辣椒抗黄瓜花叶病毒数量性状基因座的鉴定[[]。理论与应用遗传学,2001,102( 8) : 1213 - 1220。
[3]马祖里克·米,莫里亚蒂·格,格洛思·米,等.和平工作:有机系统用抗黄瓜花叶病毒的早期红甜椒[.霍特科学,2009,44 ( 5 ) : 1464 -1467。
[4]张朱庆。辣椒病毒病[研究。长沙:湖南农业大学,2009。
[5]赵娟。辣椒抗黄瓜花叶病毒分子连锁图谱的构建及QTL图谱的构建。呼和浩特:内蒙古农业大学,2009。


