血管紧张素转化酶2的结构与生物学作用,血管紧张素转换酶
血管紧张素转化酶2的结构与生物学作用
血管紧张素转换酶用于治疗心脑血管疾病,如高血压。肾素-血管紧张素-醛固酮系统在高血压的发生发展中起着重要作用,其中血管紧张素ⅱ是主要的效应肽。其临床意义在于抑制血管紧张素转换酶(ACE)对血管紧张素转换酶(RAS)的抑制能有效预防高血压的发生和发展。临床治疗
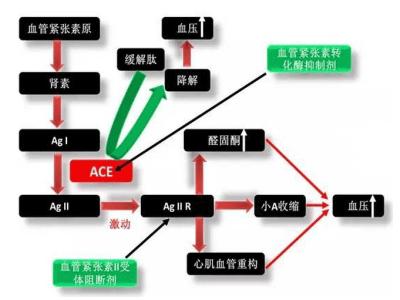
血管紧张素转换酶抵制剂的作用机制是什么?
ACEI-血管紧张素转换酶抑制剂,也称为血管紧张素转换酶抑制剂,即抗高血压多肽 ACEI,一种血管紧张素转换酶抑制剂,首次作为从蛇毒中提取的九肽tipotein在临床上应用。近年来,开发了20多种ACEI药物。目前,雷米提利等药物在临床上得到广泛应用。 除了有效降低血压,血管紧张素转换酶(血管紧张素转换酶3.4.15.1)是一种外肽酶 1功能编辑器的主要功能如下:催化血管紧张素ⅰ转化为血管紧张素ⅱ;停用缓激肽 血管紧张素转换酶抑制剂已成为治疗高血压、心力衰竭和肾小球疾病预后的独立因素。蛋白尿是影响肾小球疾病预后的独立因素。长期大量蛋白尿不仅会导致低蛋白血症,还会引起肾小球和肾小管的毒性和炎症反应。 长期高血压和肾小球内压升高都会损害肾功能,肾功能是影响各种肾小球疾病预后的重要因素。可以降低全身血压和肾小球内压。血管紧张素转换酶正常值(1)三硝基苯磺酸钠(INBS)显色方法:26.1 ~ 56.7 ku/l (2)酶偶联法:289 83U/L 血清血管紧张素转换酶的测定主要与肺部疾病的诊断相同,对其他系统性疾病的诊断和治疗也有一定的价值。 (1)肺部疾病:绝大多数结节病患者血清中血管紧张素转换酶活性升高,其正负效应明显。血管紧张素转换酶抑制剂;抑制血管紧张素转换酶的ACEI化合物 血管紧张素转换酶催化血管紧张素ⅰ生成血管紧张素ⅱ,血管紧张素ⅰ是一种强血管收缩剂,也是肾上腺皮质醛固酮释放的激活剂。 ACEI可以抑制血液的生长,
血管紧张素转换酶
血管紧张素转换酶用于治疗心脑血管疾病,如高血压。肾素-血管紧张素-醛固酮系统在高血压的发生发展中起着重要作用,其中血管紧张素ⅱ是主要的效应肽。其临床意义在于抑制血管紧张素转换酶(ACE)对血管紧张素转换酶(RAS)的抑制能有效预防高血压的发生和发展。临床治疗
血管紧张素转换酶抵制剂的作用机制是什么?
血管紧张素转化酶2的结构与生物学作用范文

血管紧张素转换酶2是肾素-血管紧张素系统的关键酶。血管紧张素转换酶2被认为是血管紧张素转换酶-血管紧张素ⅱ-血管紧张素受体ⅰ轴引起的血管收缩、增殖、纤维化和炎症的内源性调节剂。ACE2可以维持RAS系统的平衡。RAS系统中的血管紧张素转化酶(ACE)是二肽酶,能水解底物Angⅰ羧基端的两个氨基酸为Angⅱ,Angⅱ是RAS系统中的主要活性肽,Angⅱ与ATI结合产生一系列生物功能。乙酰胆碱酯酶2与乙酰胆碱酯酶具有同源性。ACE2是一种单肽酶,能从底物的羧基端解离氨基酸。其底物包括血管紧张素ⅰ、血管紧张素ⅱ、9位精氨酸缓激肽、孤儿G蛋白偶联受体(apelin) -13、内啡肽和加压素[1,2]。ACE2可将十肽Angⅰ羧基端的亮氨酸水解成九肽Ang 1 ~ 9,ACE2可将八肽Angⅱ(1 ~ 8)羧基端的苯丙氨酸水解成七肽Ang 1 ~ 7,Ang 1 ~ 7可与细胞膜上的G蛋白偶联受体团结合,从而抑制Angⅱ[3的聚集。乙酰胆碱酯酶2转化血管紧张素ⅱ为血管紧张素1 ~7的催化活性比血管紧张素ⅰ转化血管紧张素1 ~9的催化活性高400倍。
乙酰胆碱酯酶2比其他肽具有更强的催化活性,能将血管紧张素ⅱ转化为血管紧张素1 ~ 7。ACE-Angⅱ-ATI拮抗ACE2-Ang 1 ~ 7-Mas [5,6]。本文综述了ACE2的功能。
1 ACE2的结构
ACE2属于1型跨膜糖蛋白,含有805个氨基酸。ACE2主要以膜连接蛋白的形式存在,可溶性ACE 2存在于尿液和血浆中。ACE 2的胞外结构域包含具有催化活性的金属钛酶结构区(锌结合区),其与ACE的催化区有42%的序列一致性,61%的序列相似性为[4]。ACE2的主要底物是Ang,它也能与低亲和力的其他底物[5,7 ~ 9]反应。目前,已经证明ACE3、collectrin和ACE在其他哺乳动物中具有同源性。乙酰胆碱酯酶2细胞的氮端没有催化活性,其氨基酸序列与collectrin的48%相同。
ACE2可以从细胞膜上脱落,进入循环和尿液。乙酰胆碱酯酶2在血浆中的活性极低,可用作内源性抑制剂。肿瘤坏死因子转化酶(TACE/ADAM17)与ACE2 [10胞外结构域的解离有关。
ACE2的胞质尾区含有一个可与钙调素连接的位点,这与胞外区的活性催化结构区从细胞膜上解离有关。
ace2的生物学效应
ACE2是RAS系统的重要组成部分,在肾、肺、胃、肠、骨髓、脾、肝、视网膜、胎盘、卵巢、脑组织、心脏、睾丸、冠状动脉、动脉、静脉、脂肪组织、内皮细胞、巨噬细胞等组织细胞中表达。[2,7,9,11]。血管紧张素转换酶2能拮抗血管紧张素转换酶的作用,调节组织中信号肽的平衡。它在不同的疾病阶段起着重要的作用。
2.1 ACE2和肿瘤RAS系统通过肿瘤微环境中的旁分泌影响肿瘤生长、血管生成、肿瘤侵袭和转移。血管紧张素转换酶2作用于血管紧张素ⅱ,产生内源性7肽血管紧张素1 ~ 7,血管紧张素1 ~ 7结合MAS受体,通过一系列信号转导发挥抗肿瘤细胞增殖、血管生成、侵袭和转移的作用RAS系统抑制肿瘤微环境中的血管生成。Ang 1 ~ 7通过去磷酸化叉头转录因子(FOX01)的Ser256激活转录因子FOX01,FOX01转位至内皮细胞和肿瘤细胞核,激活与凋亡、细胞周期停滞和氧化应激耐受相关的基因。血管紧张素1 ~7通过激活[16,17抑制内皮细胞血管生成。一氧化氮在小鼠体内的释放与Ang 1 ~ 7 [18的抗血管生成特性有关。体外实验表明,Ang 1 ~7能抑制内皮细胞内腔的形成。Ang 1 ~7能抑制人肺癌A549细胞腔的形成,降低肿瘤内的血管密度。血管紧张素1 ~7通过降低血管内皮生长因子(血管内皮生长因子)和胎盘生长因子(PIGF)来增加可溶性血管内皮生长因子受体(sFlt) -1,从而抑制促进血管生成的信号转导途径ACE2基因敲除增加慢性肝损伤小鼠模型肝脏胶原和肿瘤坏死因子-α,加重肝纤维化。注射重组乙酰胆碱酯酶2可改善小鼠肝纤维化。在慢性肝损伤小鼠模型中,肝脏乙酰胆碱酯酶2活性增强,血浆血管紧张素1 ~7水平升高,4月亮JY。血管紧张素转换酶2和血管紧张素-( 1-7)在高血压肾中的作用〔J〕。电解质血液出版社,2011;9( 2) : 41-4。。血管紧张素1 ~7注入结扎胆道的无刷直流鼠体内,降低肝脏胶原和羟脯氨酸含量。ⅰ型胶原α1基因(胶原酶1)、α-形状记忆合金、血管内皮生长因子和结缔组织生长因子(CTGF)的表达降低,肝纤维化改善。MAS拮抗剂A799促进BLD小鼠肝脏胶原沉积、转化生长因子(TGF) -β1和羟脯氨酸的增加。血管紧张素转换酶2-血管紧张素1 ~ 7-MAS轴在肝脏中具有抗纤维化作用特纳行政区克拉克街5号。血管紧张素转换酶2:第一版〔lJ〕。国际高血压杂志,2012;2012年:307-15。。。。人肺癌细胞A549中ACE2的过表达和血管紧张素ⅱ的降低抑制了肿瘤细胞的增殖。其机制是安格1 ~ 7与MAS受体的结合能降低环氧化酶2,抑制丝裂原活化蛋白激酶信号转导通路[13,14]。血管紧张素1 ~ 7可通过减少肿瘤相关成纤维细胞的增殖和纤维化反应来减少人乳腺癌的原位体积。其机制是Ang 1 ~ 7增加MASK磷酸酶(DUSP) 1的表达,去磷酸化MASK并抑制信号转导AKT通过去磷酸化下游的FOX01 Ser 256激活FOX01,这有利于肝糖原的输出,而磷酸化的FOX01 Ser 256、Ser319、Thr24使其从细胞核进入细胞质并失去转录活性。Ang 1 ~ 7诱导PI3KC2A Ser 338磷酸化,将葡萄糖转运蛋白4转运至细胞膜。葡萄糖转运蛋白4与葡萄糖转运有关。Ang 1 ~7能调节脂肪代谢。TGR( A1 ~ 7) 3292只正常喂养的动物血液中的血管紧张素1 ~ 7增加,甘油三酯、胆固醇和脂肪组织减少。MAS消除导致FVB/北美小鼠血脂异常、胰岛素抵抗、高瘦素和低脂联素。与野生型相比,它增加了腹部脂肪组织,减少了糖的摄入[22,23]。。组蛋白去乙酰化酶(HDAC) 1在实体肿瘤中过表达,具有促进肿瘤生长的作用。血管紧张素1-7通过去磷酸化丝氨酸421和丝氨酸423抑制肿瘤增殖,HDAC1 2.3血管紧张素转换酶2(ACE2)和动脉粥样硬化在动物和人动脉粥样硬化中,血管内皮细胞、平滑肌细胞和巨噬细胞内存在ACE2的mRNA和蛋白表达,表明ACE 2的表达与动脉粥样硬化有关。动脉粥样硬化的早期事件是内皮功能障碍。ACE2的过表达可以改善内皮依赖性血管舒张,增加内皮细胞腔的形成活性,促进内皮细胞的迁移。乙酰胆碱诱导的乙酰胆碱缺乏小鼠主动脉内皮依赖性血管舒张受损。氧化应激是动脉粥样硬化的病理特征。ACE2脐静脉内皮细胞的过度表达减少了血管紧张素ⅱ诱导的活性氧自由基的形成、核因子κB的脱氧核糖核酸连接活性和p22 phoxp表达。ACE2通过Ang 1 ~ 7发挥抗感染作用。主动脉中ACE2表达的减少增加促炎因子的表达,如肿瘤坏死因子α(肿瘤坏死因子α)、白细胞介素(白细胞介素)-6、单核细胞趋化蛋白(单核细胞趋化蛋白)-1、血管细胞粘附分子(VCAM) -1、基质金属蛋白酶-2、基质金属蛋白酶-9,这些因子有助于白细胞粘附于内皮细胞和血管壁。ACE2调节巨噬细胞和内皮细胞之间的粘附。血管内皮细胞中ACE2的过度表达降低了肿瘤坏死因子-α诱导的巨噬细胞与内皮细胞之间的粘附;ACE2沉默增加粘附力。ACE2抑制血管平滑肌细胞的增殖和迁移。在体内,ACE2的过表达降低了与血管平滑肌细胞增殖相关的ERK1/2、p38激活和JAK2、信号转导和转录激活因子(STAT) 3蛋白水平。血管平滑肌细胞中过表达的乙酰胆碱酯酶2能抑制血管紧张素ⅱ诱导的基质金属蛋白酶-9的表达,从而抑制血管平滑肌细胞的迁移。ACE2可通过Ang 1 ~ 7抑制单核细胞粘附和迁移。乙酰胆碱酯酶2缺乏的人单核细胞白血病(THP-1)巨噬细胞对内皮细胞的粘附力增加。过表达的乙酰胆碱酯酶2能抑制血管紧张素ⅱ诱导的单核细胞趋化蛋白-1在THP-1巨噬细胞2贾惠普,DC,谭平,等.人气道上皮血管紧张素转换酶2的外域脱落[J].肺生理学,2009;297( 1) : L84-96。上的表达。。
[12]
血管紧张素转换酶2-血管紧张素1 ~ 7-MAS轴与肿瘤侵袭和转移有关。基质金属蛋白酶能降解细胞外基质,细胞外基质是肿瘤侵袭和转移的标志。Ang 1 ~7能抑制基质金属蛋白酶-2和基质金属蛋白酶-9的表达和活性。其基质是抑制磷酸肌醇3激酶/蛋白激酶B( PI3K/AKT)、p38和c-Jun氨基末端激酶(JNK) 2.6 ACE2和哈特尔普尔氏病哈特尔普尔氏病患者可能患有光敏皮疹、智力迟钝、烦躁、身材矮小、头痛、步态不稳、虚脱或晕厥。中性氨基酸转运蛋白(BOAT) 1基因的突变与哈特尔普尔病的发生有关。ACE2在肠上皮细胞中高度表达,并可与BOAT1中性氨基酸转运蛋白连接,从而促进氨基酸重新吸收到血液中。当氨基酸缺乏时,可能会发生肠道炎症和腹泻。ACE2可以通过氨基酸稳态、抗菌肽的表达和肠道微生物的平衡来调节肠道上皮细胞的免疫6巴特列·德,何塞·索勒·米,叶·米·阿西2与糖尿病:阿西的阿西〔日本〕糖尿病,2010;59( 12) : 2994-6。。的信号转导途径。上皮-间质转化(EMT)是上皮细胞通过特定程序转化为间质表型细胞的生物学过程,在癌症转移中起着重要作用。其主要特征包括细胞粘附分子E-钙粘蛋白表达减少,细胞角蛋白细胞骨架转化为波形蛋白为主的细胞骨架,以及间充质细胞的形态学特征。ACE2在肺癌细胞A549中的过表达增加了E-cad-herin的表达,下调了波形蛋白和α-平滑肌肌动蛋白的表达。这表明乙酰胆碱酯酶2可以通过抑制肺癌细胞的上皮间质转化来抑制肿瘤的侵袭和转移,2.7血管紧张素转化酶ACE 2)和其他血管紧张素转化酶ACE 2)产生的血管紧张素转化酶1 ~ 7,通过调节不同来源的一氧化氮释放,可减少缺血性卒中的脑梗死面积和脑功能障碍,从而起到脑保护作用。此外,脑组织中的血管紧张素1 ~7可调节学习记忆功能。Ang 1 ~7的表达和生殖系统调节子宫内膜功能、精子发生、卵泡成熟、排卵和妊娠。在兔玻璃体中注射血管紧张素1 ~7可降低眼压,提示血管紧张素1 ~7可能具有降低眼压的作用。糖尿病大鼠眼内注射血管紧张素1 ~7可减少糖尿病视网膜病变3篇参考文献。。
2.2乙酰胆碱酯酶2和代谢综合征代谢综合征的特征是高血压、肥胖、脂质代谢异常和胰岛素抵抗(高胰岛素血症,空腹部高血糖和糖耐量受损)。胰岛素抵抗是代谢综合征的中心环节,也是代谢综合征其他异常的基础。局部RAS通过自分泌和旁分泌存在于胰腺和脂肪组织中。饮食可以调节局部RAS。高糖饮食增加胰腺中血管紧张素原、血管紧张素转换酶和AT1的表达。高糖高脂饮食增加脂肪组织中乙酰胆碱酯酶2和血管紧张素1 ~7的表达。血管紧张素转换酶抑制剂和AT1拮抗剂可以改善高血糖症。乙酰胆碱酯酶2的上调可改善糖尿病小鼠高血糖。目前,已证实RAS系统中的Ang 1 ~7可通过糖摄入和抗胰岛素抵抗相关的氧化应激改善代谢综合征。Ang 1 ~7和胰岛素在人血管内皮细胞和小鼠心脏中具有一些相同的信号转导效应酶。Ang 1 ~7可通过人血管内皮细胞MAS受体磷酸化PI3K/AKT激活信号转导通路。Ang 1 ~ 7去磷酸化非受体酪氨酸激酶c-Src和细胞外信号调节激酶(ERK1/2),抑制烟酰胺腺嘌呤二核苷磷酸氧化酶活性,负调节Angⅱ/AT1信号转导功能。Ang 1 ~7可通过小鼠心脏胰岛素受体底物(IRS-1)和Janus激酶2 (JAK2)的AT1磷酸化,调节下游以下八种胰岛素信号转导中效应酶的磷酸化:AKT1( RAC-α丝氨酸/苏氨酸蛋白激酶)、PRAS40/AKT1S1(富含脯氨酸、分子量为40kd/富含脯氨酸的AKT底物蛋白AKT底物蛋白1、荚膜蛋白(CAV1)、FOXO1、MAPK1/ERK2、paxn、PI3KC2A、vim。ang 1-7诱导一类PI3K活化。PI3K磷酸化苏氨酸(Thr) 308、丝氨酸473和苏氨酸124。活化的AKT1可以磷酸化PRAS40以解离PRAS40和雷帕霉素靶蛋白(mTOR)的复合物。mTOR的激活与胰岛素抵抗有关,而PRAS40、PRAS40和mTOR复合物的去磷酸化共同抑制mTOR活性。
[15]
[16]
2.血管紧张素转化酶2和血栓血管紧张素转化酶2-血管紧张素转化酶1 ~ 7-MAS轴具有抗血栓形成作用。乙酰胆碱酯酶2通过水解产生血管紧张素1 ~ 7,血管紧张素1 ~ 7与MAS受体结合抑制血栓形成。活化血小板的粘附、聚集和释放与血栓形成有关。马斯。血管紧张素1 ~7受体存在于内皮细胞和血小板上,可通过内皮细胞抑制血小板活化。Ang 1 ~7与内皮细胞上的MAS受体结合,通过PI3K/Akt/内皮一氧化氮合酶信号转导途径刺激前列环素的产生和一氧化氮的释放。一氧化氮可以增加血小板中的环磷尿苷,抑制血小板活化,而环磷尿苷与血小板受体结合,增加血小板中的环磷腺苷,抑制血小板活化。Ang 1 ~7可直接抑制血小板活化。Ang 1 ~ 7与血小板上的MAS受体结合,增加血小板中一氧化氮的合成,抑制血小板的活化。3魏索基,叶明,罗德里格斯。用重组血管紧张素转换酶2靶向血管紧张素ⅱ降解:血管紧张素ⅱ依赖性高血压的预防〔J〕。高血压,2010;55( 1) : 90-8。。
2.5乙酰胆碱酯酶2和肝纤维化乙酰胆碱酯酶2-血管紧张素1 ~ 7-MAS轴在肝、肺、心、肾等不同器官中具有抗纤维化作用。肝纤维化是由多种原因引起的慢性肝损伤引起的病理变化,表现为细胞外基质成分(尤其是胶原蛋白)在肝脏中过度异常沉积,影响肝功能。乙酰胆碱酯酶2通过降解血管紧张素ⅱ和产生血管紧张素1 ~ 7抑制肝纤维化。
[19]
[20]
[21]
[22]
1布拉德福中国,伊利博士,雷扎达MK .靶向肾素-血管紧张素系统的血管保护轴:肺高血压治疗的一种新的战略方法[J].当前高血压代表,2010;12( 4) : 212-9。
[24]
[25]
[26]
[27]
[28]
7托马斯·麦卡利斯·C。血管紧张素转换酶2 ( ACE2)是肾素血管紧张素系统在健康和疾病中的一个重要调节因子[J].国际消化杂志,2012;2012年:256-94。
8申奥五,齐晔,卡托维奇·梅杰,等.血管紧张素转换酶2,肺动脉高压的一个有希望的治疗靶点〔J〕.《柯尔平药理》,2011年;11 ( 2 ) :150-5。
《乙酰胆碱酯酶2的甜的一面:在糖尿病中起作用的生理学证据》〔期刊〕摩尔细胞内分泌学,2009;302 ( 2 ) :193-202。
贾利勒·JE,奥卡兰扎议员。血管紧张素转换酶2/血管-( 1-9)轴在血管重塑中的保护作用〔J〕.国际高血压杂志,2012;2012年:594361。
夏海,拉扎提格斯。脑中血管紧张素转换酶2:性质和未来方向〔J〕。神经化学杂志,2008;107( 6) : 1482-94。
12帕索斯-席尔瓦DG,维拉诺-布拉加特,桑托斯拉。血管紧张素-( 1-7):超越心肾作用〔J〕。临床科学(伦敦),2013;124( 7) : 443-56。
塔兰特行政区加拉格尔广场13号。血管紧张素-( 1-7)对人肺癌细胞生长的抑制25( 11) : 2045-52。
14梅农·杰,索托-潘托贾博士,卡拉汉·MF,等.血管紧张素-( 1-7)抑制人肺腺癌裸鼠移植瘤的生长,通过环氧化酶-2的减少[J].癌症研究,2007;67( 6) :2809-15。
15库克KL,Metheny-Barlow LJ,Tallant EA,等.血管紧张素-( 1-7)减少原位乳腺肿瘤纤维化[J].癌症研究,2010;70( 21) : 8319-28。


